DNA微阵列技术:计算序列设计方法
需积分: 5 173 浏览量
更新于2024-07-18
收藏 973KB PDF 举报
"DNA微阵列科技"
DNA微阵列是一种生物技术,用于同时检测大量DNA序列的表达水平或存在情况。这项技术的核心是利用DNA序列的互补性原理,通过将成千上万的小片DNA固定在固体支持物(如玻璃片、芯片)上形成微阵列,每个小片代表一个特定的DNA序列,即探针。当样本DNA与微阵列接触时,如果样本中存在与探针互补的序列,两者会通过氢键结合,形成杂交分子,从而被检测到。
SLS算法(Shortest Linear Superstring Problem)在DNA微阵列设计中扮演重要角色。这个算法主要用于构建种子序列,即较短的DNA片段,它们是构建更长序列的基础。SLS的目标是找到一系列短串,使它们能够通过重叠连接形成最长的线性串。在DNA微阵列设计中,这些种子序列用于覆盖目标基因组区域,确保能捕获所有感兴趣的序列信息。
团搜索算法是另一种用于DNA序列分析的方法。在微阵列设计中,它可能用于寻找具有特定属性(如高度保守区域或特异性序列)的DNA片段集合。这种算法可以优化探针的选择,提高检测的特异性和敏感性。
构建种子的方法涉及选择具有适当长度、特异性和覆盖度的DNA序列。种子序列的长度通常在15-25个碱基对之间,过短可能导致匹配不准确,过长则可能增加成本。特异性是指种子序列只应与目标序列完全匹配,避免与其他非目标序列发生交叉反应。覆盖度则确保微阵列中的探针能够覆盖整个感兴趣的基因区域。
DNA编码则是利用DNA的四种核苷酸(腺嘌呤A、胸腺嘧啶T、胞嘧啶C和鸟嘌呤G)作为信息存储和处理的介质。在DNA微阵列中,编码技术可能用于标记探针或样本,以便区分不同的序列和后续的分析步骤。例如,可以通过特定的DNA序列标签来标识不同的样本来源或实验条件。
DNA微阵列科技结合了生物信息学和计算系统生物学的方法,如SLS算法、团搜索算法和DNA编码,以高效地设计和使用微阵列进行大规模的基因表达分析或遗传变异检测。这些技术的不断发展和优化,对于理解复杂的生物过程、疾病机制以及药物研发具有重大意义。
5
uses a traditional approach. The criteria taken into account are specificity (use of BLAST), melting temperature and
position in the transcripts. ProMide (Rahmann, 2003) consists of a set of Perl scripts, with a C program for basic
calculations, which uses the longest common substring as a specificity measure for the oligonucleotides. It uses
complex data structures such as ‗enhanced suffix array‘ and some statistical properties of sequences. Oliz (Chen &
Sharp, 2002) is also implemented in Perl and uses a classical approach relying on BLAST for specificity testing. It
however requires additional software to function, like cap3, clustalw, EMBOSS ―prima‖ and a database in the UniGene
format. The originality of the method stems from the fact that the oligonucleotides are searched in the 3‘ untranslated
region of the mRNAs, a very specific area where sequences are largely available through expressed sequence tag
projects.
The majority of the client-server and autonomous software described above use a pipelined filtering approach for the
design of microarray probes, where at an initial stage a local alignment approach is employed for cross-hybridization
avoidance and then each ulterior filter evaluates the quality of each probe and ranks them according to well defined
scoring criteria.
DNA SEQUENCE DESIGN - PROBLEM DESCRIPTION
A single stranded DNA molecule is a long, unbranched polymer composed of only four types of subunits. These
subunits are the deoxyribonucleotides containing the bases adenine (A), cytosine (C), guanine (G) and thymine (T). The
nucleotides are linked together by chemical bonds and they are attached to a sugar-phosphate chain like four kinds of
beads strung on a necklace (Figure 1). The sugar-phosphate chain is composed by alternating the ribose and phosphate
for each nucleotide. This alternating structure gives each base and implicitly the whole strand a direction from the
ribose end (denoted by 5‘) to the phosphate end (denoted by 3‘). When the 5‘/3‘ ends are not explicitly labeled, DNA
sequences are assumed to be written in 5‘ to 3‘ direction.
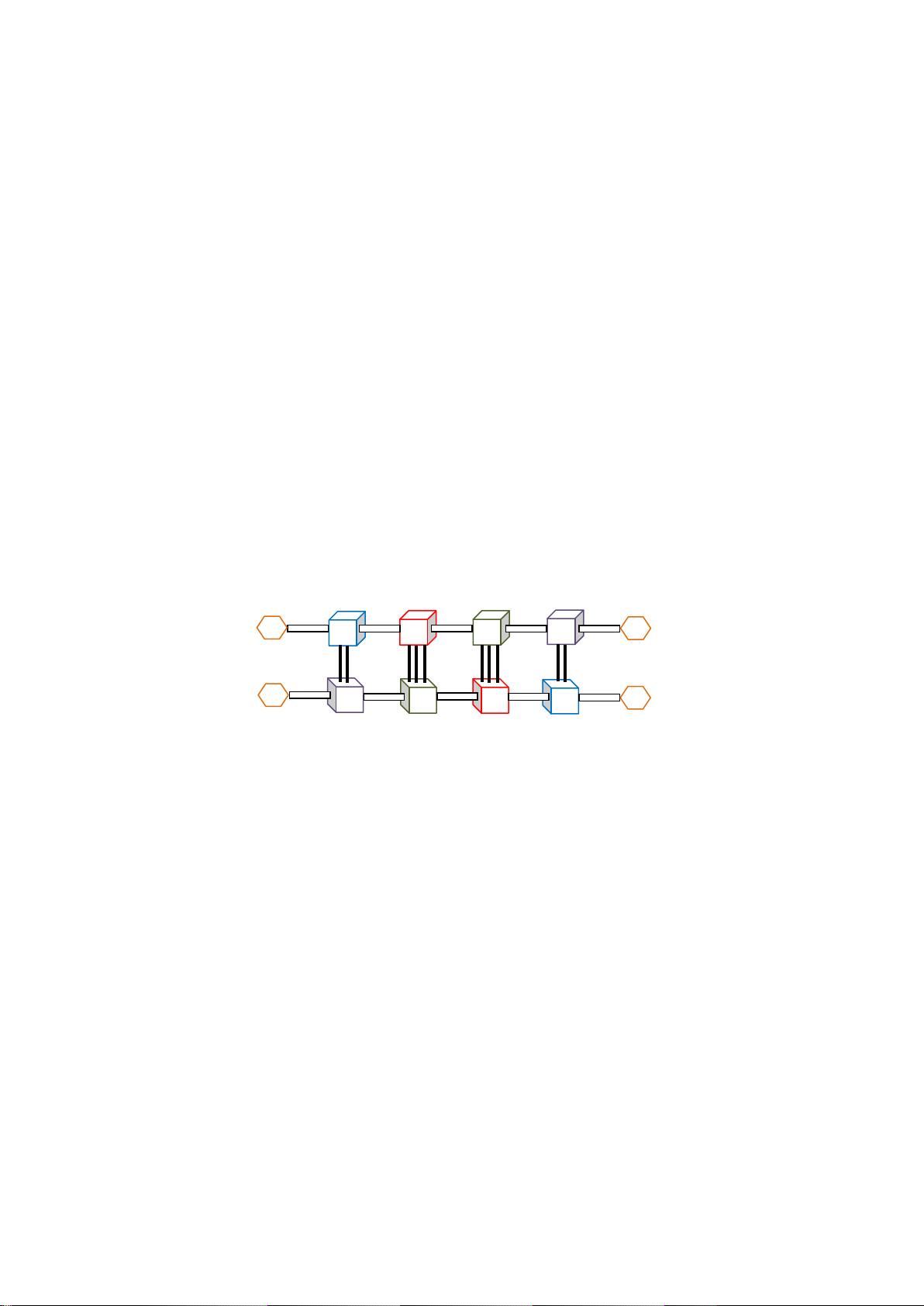
Figure 1: Idealized structure of a pair of hybridized DNA sequences.
The nucleotides composing DNA bind to each other in pairs via hydrogen bonds in a process known as hybridization.
Each nucleotide pairs up with its unique complement (the Watson-Crick complement), so C pairs up with G and A with
T. C and G pair up in a more stable manner than A and T do, which is due to one extra hydrogen bond in the C-G pair.
The Watson-Crick complement of a DNA strand is the strand obtained by replacing each C nucleotide with a G and
vice versa, and each T nucleotide with an A and vice versa, and also switching the 5‘ and 3‘ ends. For example the
Watson-Crick complement of 5‘-AACTAG-3‘ is 3‘-TTGATC-5‘. Non-Watson-Crick base pairings are also possible.
For example, G-T pairs occur naturally in biological sequences (Ho et al., 1985; Pfaff et al., 2008).
Formally, these notions are captured in the following definitions:
A DNA strand w is represented by a string over the quaternary alphabet {A, C, G, T}. String w corresponds to a single-
stranded DNA molecule with the left end of the string corresponding to the 5‘-end of the DNA strand.
The complement, or Watson-Crick complement, of a DNA strand w is obtained by reversing w and then by replacing
each A with a T and vice versa, and replacing each C in the strand by G and vice versa. We denote the Watson-Crick
complement of a DNA strand w with c(w) and we will simply use the terms complementary strand, or complement in
this work. For simplicity, we simply use c
i
instead of c(w
i
).
Throughout the remainder of the chapter we use symbols w
1
, w
2
, ..., w
k
to denote unique strands and c
1
, c
2
, ..., c
k
to
C
A
G
T
C
A
G
T

剩余30页未读,继续阅读
2021-07-26 上传
2021-07-26 上传
2021-07-26 上传
2021-07-26 上传
2022-04-10 上传
2021-07-26 上传
2021-09-25 上传
2021-11-22 上传
2022-04-10 上传

凤凰AI
- 粉丝: 224
- 资源: 391
 我的内容管理
展开
我的内容管理
展开







最新资源
- 探索AVL树算法:以Faculdade Senac Porto Alegre实践为例
- 小学语文教学新工具:创新黑板设计解析
- Minecraft服务器管理新插件ServerForms发布
- MATLAB基因网络模型代码实现及开源分享
- 全方位技术项目源码合集:***报名系统
- Phalcon框架实战案例分析
- MATLAB与Python结合实现短期电力负荷预测的DAT300项目解析
- 市场营销教学专用查询装置设计方案
- 随身WiFi高通210 MS8909设备的Root引导文件破解攻略
- 实现服务器端级联:modella与leveldb适配器的应用
- Oracle Linux安装必备依赖包清单与步骤
- Shyer项目:寻找喜欢的聊天伙伴
- MEAN堆栈入门项目: postings-app
- 在线WPS办公功能全接触及应用示例
- 新型带储订盒订书机设计文档
- VB多媒体教学演示系统源代码及技术项目资源大全
