基因递送与表达调控:混合载体设计的进展与策略
需积分: 5 87 浏览量
更新于2024-08-07
收藏 341KB PDF 举报
本文是一篇学术论文,标题为“Hybrid vector designs to control the delivery, fate, and expression of transgenes”,由Paula Y. P. Lam和Xandra O. Breake撰写。两位作者分别来自美国马萨诸塞州总医院分子神经遗传学单元以及哈佛医学院神经科学项目,以及新加坡国家癌症中心细胞与分子研究分部。论文主要关注的是基因疗法中的关键挑战——如何精确地将基因(transgenes)送至疾病发生部位,并调控其表达,同时避免副作用。
病毒载体设计在基因疗法中起着至关重要的作用。早期的研究主要集中在各种复制缺陷型病毒(replication-defective viral mutants)上,这些病毒在宿主细胞内不能自我复制,但仍然能携带和释放基因。随着技术的发展,研究人员逐渐转向复制条件依赖型病毒(replication-conditioned viruses),这些病毒在特定条件下才会激活并进行复制,从而增加基因表达的精确性。
论文指出,理想的混合病毒设计(hybrid vectors)旨在结合多种优势,包括但不限于:提高病毒对目标细胞的选择性,确保基因能在目标组织特异性地定位;通过调控病毒的复制周期或表达水平,实现精细的基因表达控制;以及减少病毒的免疫反应,降低副作用。此外,文章可能还探讨了非病毒载体系统,如使用质粒、微粒体、纳米粒子等,作为替代方案,以减少潜在的免疫排斥和安全性问题。
论文作者强调,要实现基因疗法的成功,关键在于持续优化和创新病毒载体设计,这不仅涉及病毒本身的特性改良,还包括对宿主细胞生物学的理解和应用,以及对递送策略的深入研究。通过跨学科的合作,包括分子生物学、遗传学、生物工程等领域,科学家们正在努力解决这些挑战,以期为基因治疗提供更为精准、安全的工具。
这篇论文深入讨论了当前在基因疗法中使用混合病毒设计控制基因传递、命运和表达的最新进展,以及未来可能的研究方向。这对于理解基因治疗领域的前沿进展,以及推动相关技术的实际应用具有重要意义。
mediate binding to the cell surface and hence determine
cell speci®city [15,16]. The envelope fuses with the cell
membrane releasing the virion contents into the cyto-
plasm. A discrete number of different host cell surface
proteins can serve as receptors for the retroviral envelope
proteins. Receptors, such as CD4 which binds lentivirus,
are expressed by cells of speci®c lineages such as
lymphocytes, while amphotropic, xenotropic and gibbon
ape leukemia virus (GALV) receptors which bind MMLV
are expressed on most, if not all, mammalian cells.
Retroviral virions that express different envelope pro-
teins, such as VSV-G, have increased stability as well as an
expanded host range [17]. This is primarily due to the
ability of the VSV-G to recognize membrane phospholipid
as a minimal receptor [18,19]. Unlike standard murine
retroviruses, VSV-G retrovirus vectors are also relatively
resistant to deactivation by human complement [20], can
be concentrated to high titers by centrifugation, and
frozen/thawed without loss of infectivity [17]. Viral
tropism has also been altered by generating a protease-
cleavable, receptor-blocking domain onto the viral coat
protein. Exposure to a speci®c protease that can cleave
the site is suf®cient to restore virus infectivity at sites with
high local activity of the relevant protease [21]. Other
means used to target retrovirus vectors to speci®c cell
types include: chemical modi®cation, use of antibody
ligands, and creation of recombinant envelope proteins
(for a review see [22]).
Receptor-mediated entry of adenovirus into cells is
dependent on two of its coat proteins, the ®ber and
penton base. The ®ber mediates primary attachment to a
wide range of cell types via the Coxsackie-adenovirus
receptor (CAR) protein [23]. Following attachment, an
Arg-Gly-Asp (RGD) tripeptide motif in the penton base
protein binds to a
v
integrins on the host cell membrane,
which then trigger cellular internalization by receptor-
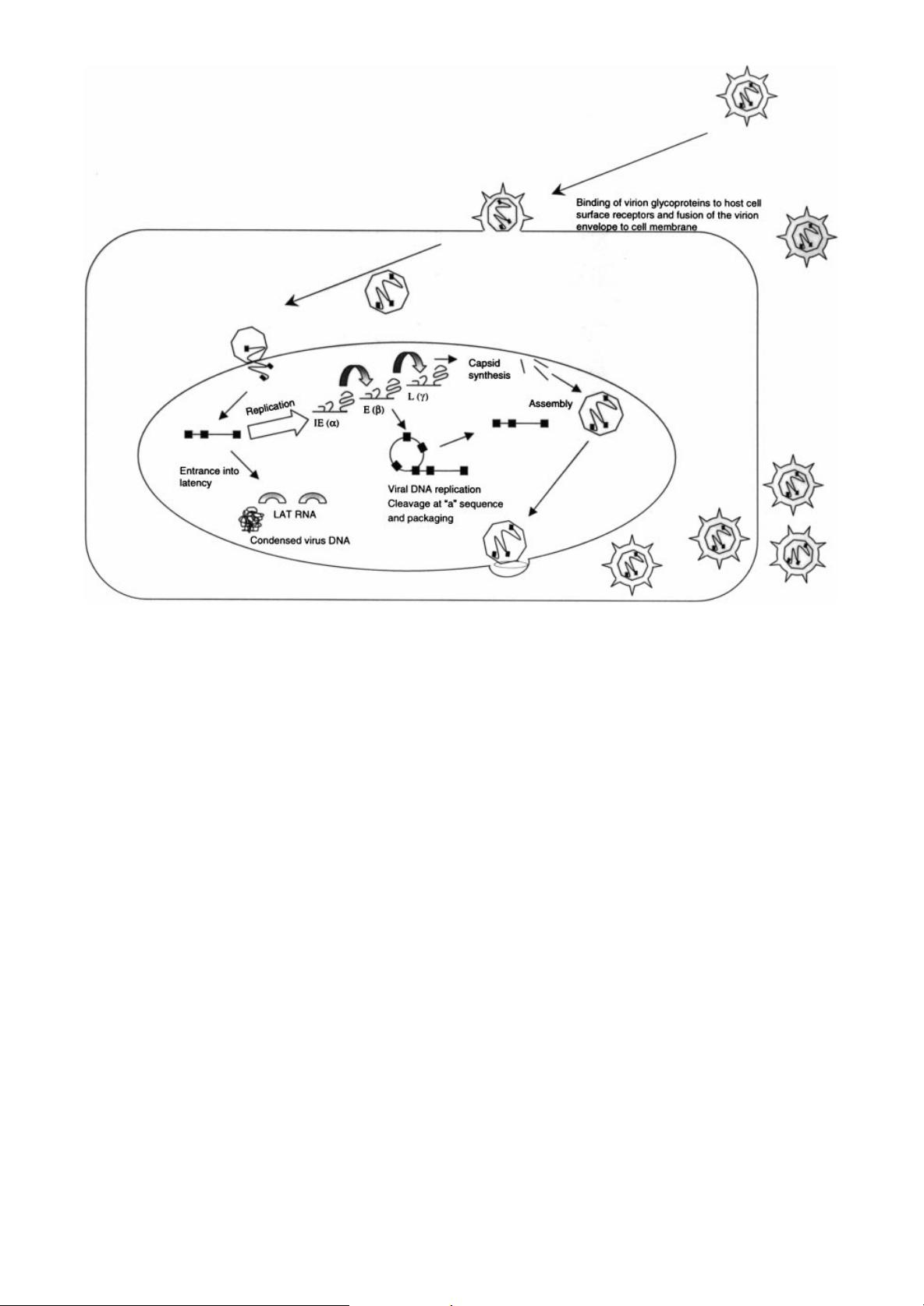
Figure 1. Schematic view of HSV infection. Glycoproteins in the envelope of the virion mediate binding to the cell surface and
fusion with the plasma membrane. The capsid containing the viral DNA is transported to the nuclear membrane via an active
microtubular mechanism. The capsid opens and extrudes the virus DNA into the nucleus through nuclear pores. Within the
nucleus, wild-type HSV can either replicate to produce more virus particles or enter latency. Virus replication involves a cas-
cade of virus gene expression involving sequential transcription of immediate early (IE), early (E) and late (L) genes. Viral
DNA is replicated as a continuum and enters newly formed, empty capsids via binding of the `a' or pac sequence. When the
capsid is full the virus DNA is cut at the pac sequence and the capsid seals closed. Capsids acquire an envelope via transport
through the nuclear and plasma membranes (for a review see [140]). Ultimately production of HSV virions leads to cell death.
Under some conditions, e.g. low MOI in sensory neurons, the virus DNA enters an episomal state of latency in the cell nucleus
characterized by a tight con®guration and production of latency associated transcripts (LATs) which is benign, but from which
the virus can reactivate to productive replication
Hybrid Viral Vector Designs 397
Copyright # 2000 John Wiley & Sons, Ltd. J Gene Med 2000; 2: 395±408.
剩余13页未读,继续阅读
2021-06-29 上传
2024-09-12 上传
2024-09-12 上传
2024-09-12 上传
2024-09-12 上传
2024-09-12 上传
2024-09-12 上传
2024-09-12 上传

weixin_38635684
- 粉丝: 7
- 资源: 955
 我的内容管理
展开
我的内容管理
展开







最新资源
- 最优条件下三次B样条小波边缘检测算子研究
- 深入解析:wav文件格式结构
- JIRA系统配置指南:代理与SSL设置
- 入门必备:电阻电容识别全解析
- U盘制作启动盘:详细教程解决无光驱装系统难题
- Eclipse快捷键大全:提升开发效率的必备秘籍
- C++ Primer Plus中文版:深入学习C++编程必备
- Eclipse常用快捷键汇总与操作指南
- JavaScript作用域解析与面向对象基础
- 软通动力Java笔试题解析
- 自定义标签配置与使用指南
- Android Intent深度解析:组件通信与广播机制
- 增强MyEclipse代码提示功能设置教程
- x86下VMware环境中Openwrt编译与LuCI集成指南
- S3C2440A嵌入式终端电源管理系统设计探讨
- Intel DTCP-IP技术在数字家庭中的内容保护
