多模态单细胞追踪技术揭示肿瘤演进的精细进化树
72 浏览量
更新于2024-06-18
收藏 2.22MB PDF 举报
"肿瘤演变的多模态单细胞追踪技术揭示出详细的进化树"
这篇文章深入探讨了一种创新的多模态单细胞追踪技术,用于研究肿瘤的演变过程。该技术结合了全基因组测序和谱系追踪,使得科学家能够以前所未有的细节水平解析肿瘤细胞的遗传变化和进化路径。通过这种方法,研究者们能够重建肿瘤的复杂进化树,揭示不同细胞群体之间的关系和演变历程。
在文章中,研究人员应用这项技术分析了结肠癌类器官,这是一种体外培养的肿瘤模型,可以模拟体内肿瘤的行为。他们发现,通过同时测量单细胞的拷贝数变异(CNV)、单核苷酸变异(SNV)以及病毒谱系条形码,可以构建出高度详细的进化树。这些数据揭示了肿瘤细胞在进化过程中经历的染色体丢失事件,例如染色体18和4的顺序丢失,这些丢失可能与肿瘤细胞的适应性优势有关。
这一发现对于理解肿瘤的异质性和耐药性形成机制具有重要意义。染色体的丢失可能导致基因的失调,进而影响肿瘤的生长、侵袭和治疗反应。通过对这些事件的精确追踪,研究者可能能够预测肿瘤对治疗的反应,并为个性化治疗策略提供依据。
文章作者Lennart Kester、Buysde Barbanson、Anna Lyubimova等人的工作展示了多模态单细胞分析在肿瘤研究领域的潜力。他们的研究不仅提供了关于结直肠癌的新见解,也为未来研究其他类型的癌症提供了有力工具和技术平台。
通过开放获取的方式发布,这篇2022年的文章《肿瘤演变的多模态单细胞追踪技术揭示出详细的进化树》鼓励了更广泛的科学社区参与讨论和进一步的研究,以期推动肿瘤生物学和精准医学的发展。通过这样的研究,我们可以期望在未来找到更有效的方法来对抗癌症,提高患者的生存率和生活质量。
Cell Genomics2,100096,2022年2月9日
3
会
开放获取
文章
基于NLA-III限制性内切酶的技术可以精确定量每个单细胞的
CNV谱。例如,我们观察到多个独特的CNV影响染色体18,这
不能通过批量WGS检测到(图2D;例如,拷贝数状态2、6和
11).与单细胞NLA-III测序平行,我们在
体外
进化的第一天和最
后一天进行了标准批量WGS。在这些批量数据中,我们在实验
结束时在重复1中观察到完整的D4然而,B等位基因频率
(BAF)显示两种等位基因仍然存在,尽管数量不等(图
S5)。这表明一部分细胞丢失了4号染色体的一个等位基因,其
余细胞丢失了另一个等位基因。为了在单细胞中证实这一点,我
们首先获得了4号染色体的二倍型,该二倍型是基于来自同一供
体的另一个类器官系,该供体完全丢失了4号染色体的一个等位
基因。然后使用该二倍型来评估每个单细胞中4号染色体的哪个
等位基因(如果有的话)丢失。事实上,在重复1中,我们观察
到314个单细胞具有D4等位基因A,96个细胞具有D4等位基因
B.也可获得18号染色体的双倍型。在这里,我们观察到18号染
色体上的所有独特缺失都涉及相同的等位基因。观察到的4号和
18号染色体的单细胞BAF是三峰的,峰值在0、0.5和1附近,表
明细胞是二倍体而不是四倍体(图S5B)。这些观察结果表明,
单细胞NLA-III测序和WGS的组合允许在单细胞中进行等位基因
特异性CNV检测。在类器官培养物中观察到的大多数缺失和扩增
也经常在CRC患者中观察到,这强调了类器官模型与研究结直
肠癌的相关性(图2C)。
我们总共在1,641个单细胞中鉴定了25个独特的CNV,具有
52个独特的CNV状态(我们将CNV状态定义为由至少两个单细
胞共享的全基因组CNV谱;STAR方法),大小范围从434个细胞
到3个细胞(图2D)。在重复1中,我们观察到具有D4和D18的
细胞的大量扩增,而在第二(重复2)和第三(重复3)实验中,
我们观察到具有D8p的细胞的扩增
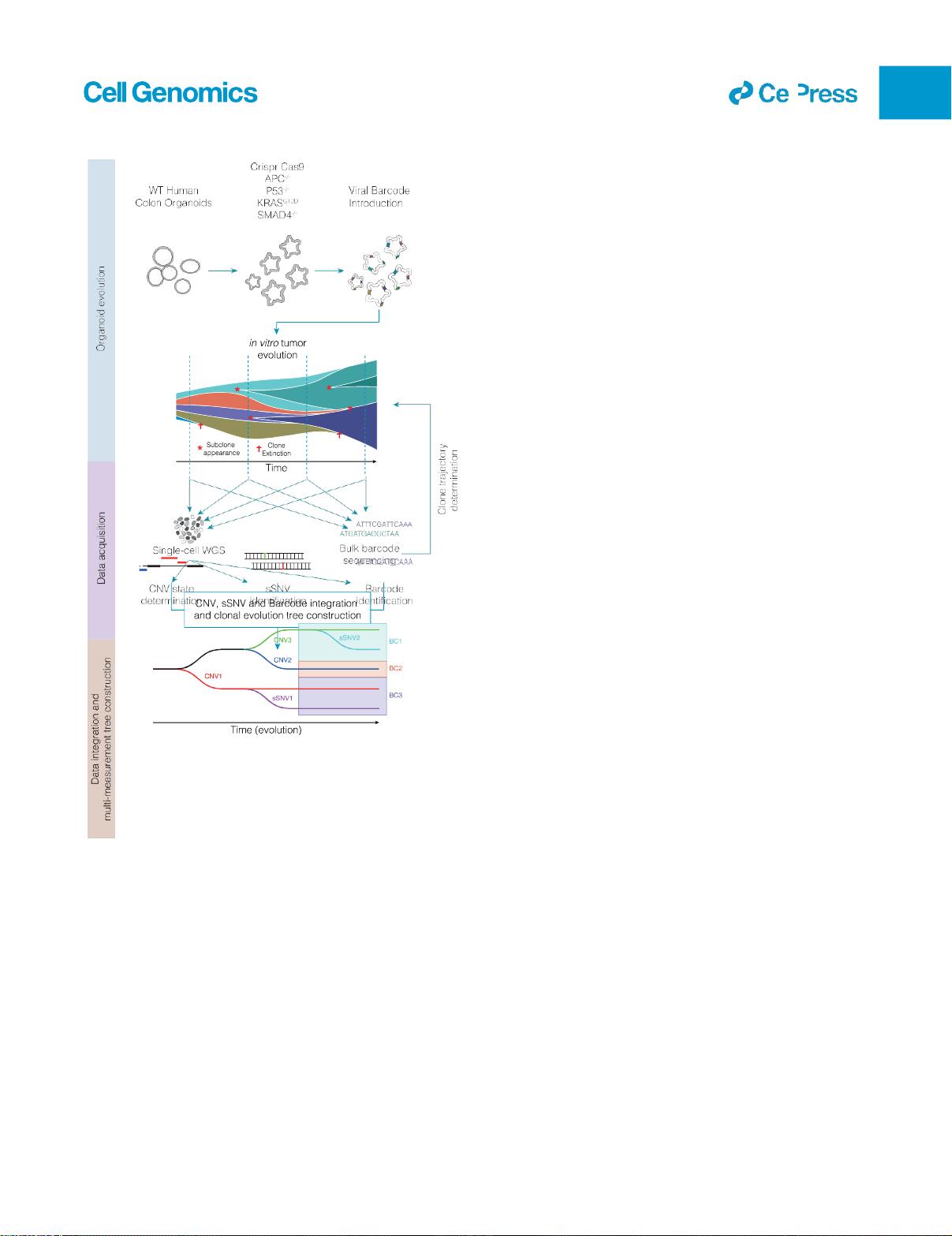
图1. 实验装置
使用基于CRISPR-Cas9的策略转化野生型人结肠直肠类器官。用引入谱系条
形码的慢病毒文库转导
22
个
转化的类器官。类器官经历了26周的
体外
进化期,
在此期间,定期进行单细胞WGS,每周评估培养复杂性。从单细胞WGS,获
得拷贝数状态、sSNV状态和谱系条形码,从而允许构建和验证高度详细的克
隆进化树。
再乘以2对中位数归一化均值使用主成分分析的离散性降低表
明,细胞通过重复和时间点聚集,而对于早期时间点,各种重复
的细胞更相似(图S3)。不同拷贝数区域之间的重复断裂点通
过分层聚类然后循环二进制分割(STAR方法)检测。的高分辨
率和低噪声(图S4)
高分辨率克隆进化树
为了构建基于CNV的初始克隆进化树,我们使用CNV状态来创
建有向编辑距离图。由于相同的CNV状态可能存在于多个时间
点,因此将每个时间点添加为图中的单独节点。这使得能够实施
时间一致性(即,较早的时间点不能从较晚的时间点导出)。用
Edmonds算法从有向CNV编辑距离图中生成生成树形图. 使用
ToverBoom可视化克隆进化树(图3C所得到的克隆进化树指示
肿瘤沿着其进化的最可能的例如,复制1的树指示CNV状态3是
CNV状态2的后代,考虑到CNV状态2具有D18并且CNV状态3具
有D18和D4,这是合乎逻辑的(图2D)。总之,高分辨率CNV
调用允许构建具有时间分量的详细克隆进化树。
剩余19页未读,继续阅读
2024-11-16 上传
2022-12-26 上传
2016-06-18 上传
2024-08-07 上传
2019-08-27 上传
2024-05-01 上传

cpongm
- 粉丝: 5
- 资源: 2万+
 我的内容管理
展开
我的内容管理
展开







最新资源
- forward_algorithm.zip_matlab例程_matlab_
- solrium:Solr的通用R接口
- newunobet:大pp
- project_euler:这是来自https的已解决问题的存储库
- webchem:来自网络的化学信息
- cartified:一个非常基本的购物车实施
- 7Applied-multi-dimensional-fusion-.zip_图形图像处理_PDF_
- risitas-uikit
- homework4-february-20-2021:Web API:代码测验
- astrofox:Astrofox是一种运动图形程序,可让您将音频转换为出色的视频
- SpotipyProject
- tdd-blog:只是学习TDD的一个示例
- ezknitr:使用“ knitr”时避免典型的工作目录痛苦
- webPass-crx插件
- vue+node少儿编程项目.zip
- test-workflow
