生物启发的深度学习:脉冲神经网络的挑战与进展
需积分: 0 133 浏览量
更新于2024-06-22
收藏 1.04MB PDF 举报
"本次综述文章探讨了脉冲神经网络(SNN)和生物启发式监督深度学习在人工智能领域的应用,旨在提供一个全面的视角来理解这两种方法的最新进展和挑战。作者首先介绍了生物神经元的基本计算原理和突触可塑性,然后深入到SNN模型的细节,讨论了其在训练过程中的独特问题。此外,文章还讨论了替代传统反向传播的生物启发式训练策略,以及如何通过生物启发的深度学习(BIDL)提升模型的计算性能和生物可信度。"
在人工智能领域,生物神经科学的研究为计算机科学家提供了丰富的灵感。脉冲神经网络(SNN)是模拟大脑神经元活动的一种模型,它以脉冲(或尖峰)的形式进行信息传递,更加接近生物神经系统的运作方式。SNN的优势在于其高效的能量消耗和时间编码能力,但与传统的基于连续数值的神经网络不同,SNN的训练通常需要解决反向传播不适用的问题。这导致了对新的训练方法的需求,例如那些受到生物过程启发的策略。
文章详细介绍了SNN模型的不同方面,包括它们如何模拟神经元的电生理特性,如 leaky integrate-and-fire (LIF) 模型,以及它们如何模拟突触权重的动态变化。在SNN的训练过程中,由于尖峰的时间序列和非连续性,传统的反向传播算法无法直接应用。因此,研究人员提出了各种生物启发式的训练方法,这些方法试图模拟生物学习规则,如 STDP(突触时间依赖性可塑性),在SNN中实现权重更新。
生物启发的深度学习(BIDL)方法是针对这一挑战的解决方案之一。这些方法不仅关注模型的性能,还注重其生物现实主义,即模型的行为和学习机制应尽可能地接近生物神经系统。BIDL试图在保持高性能的同时,提高模型的生物可信度,从而推动AI技术向更真实的神经计算模式靠拢。
总结来说,这篇综述文章提供了关于SNN和BIDL的深入洞察,涵盖了这些领域的最新研究进展和未来可能的发展方向。对于那些对生物启发式AI方法感兴趣的读者,这篇文章提供了一个宝贵的资源,帮助他们理解和利用这些技术来改进现有的神经网络模型和算法。

6 Lagani, et al.
Note that, more in general, local learning rules with higher order interactions can be considered. A general form for
such a local synaptic update equation, for a generic synaptic connection 𝑖, can be expressed as [61]:
Δ𝑤
𝑖
= 𝑎
0
+ 𝑎
1
𝑥
𝑖
+ 𝑎
2
𝑦 + 𝑎
3
𝑥
𝑖
𝑦 + 𝑎
4
𝑥
2
𝑖
+ 𝑎
5
𝑦
2
+ ... (4)
where the coecients 𝑎
𝑖
may depend on the weights.
When multiple neurons are involved in a complex network, it is desirable that dierent neurons learn to recognize
dierent patterns. In other words, neural activity should pursue a decorrelated form of coding [
51
,
154
], in order to
maximize the amount of information that neurons can transmit. This can be achieved by leveraging the biological
mechanisms of lateral and inhibitory interaction. For example, Eq. 3 can be coupled with competitive learning mechanisms
such as Winner-Takes-All (WTA) [
67
,
178
]: in this case, the neuron with strongest similarity to a given input is chosen
as the winner, and it is the only one allowed to perform a weight update, by virtue of lateral inhibition [
55
]. In this way,
if a similar input will be presented again in the future, the same neuron will be more likely to win again, and dierent
neurons will specialize on dierent clusters. Other forms of lateral interaction can also enable a group of neurons to
extract further principal components form the input (subspace learning) [15, 93, 180].
As a further extension of PCA-based learning methods, Independent Component Analysis (ICA) aims at removing
higher-order correlations from data [
3
,
17
,
74
,
85
,
91
,
162
]. Sparse Coding (SC) [
154
] is another principle, strictly
related to ICA [
153
], from which biologically plausible HaH feature learning networks can be derived, as in the Locally
Competitive Algorithm (LCA) [175].
Hebbian approaches based on competitive [
4
,
99
,
102
,
108
,
137
,
141
,
207
] or subspace learning [
10
,
86
,
104
,
109
] have
recently been subject of signicant interest towards applications in DL contexts.
4 SPIKING NEURAL NETWORKS
This Section discusses models of neural computation based on Spiking Neural Networks (SNNs) [
61
], which more
faithfully resemble real neurons compared to traditional ANN models. We start by introducing the various neuron
models for SNN simulation. We highlight the applications related to biological and neuromorphic computing, which are
of strong practical interest thanks to the energy eciency of the underlying computing paradigm, and we discuss the
challenges related to SNN traning. We describe the biological plasticity models for spiking neurons, and the approaches
to translate backprop-based training to the spiking setting. Finally, an overview of some existing experimental results is
given.
In the following, subsection 4.1 illustrates the main models of spiking neurons and spike-based strategies for
information encoding; subsection 4.2 highlights promising applications of spiking models in the eld of neuromorphic
and biological computing, and the challenges of learning with spiking models; therefore, subsection 4.3 discusses the
plasticity models for spiking neurons, based on Spike Time Dependent Plasticity (STDP); nally, subsection 4.4 presents
some experimental results from literature regarding the applications of SNNs and STDP in DL contexts.
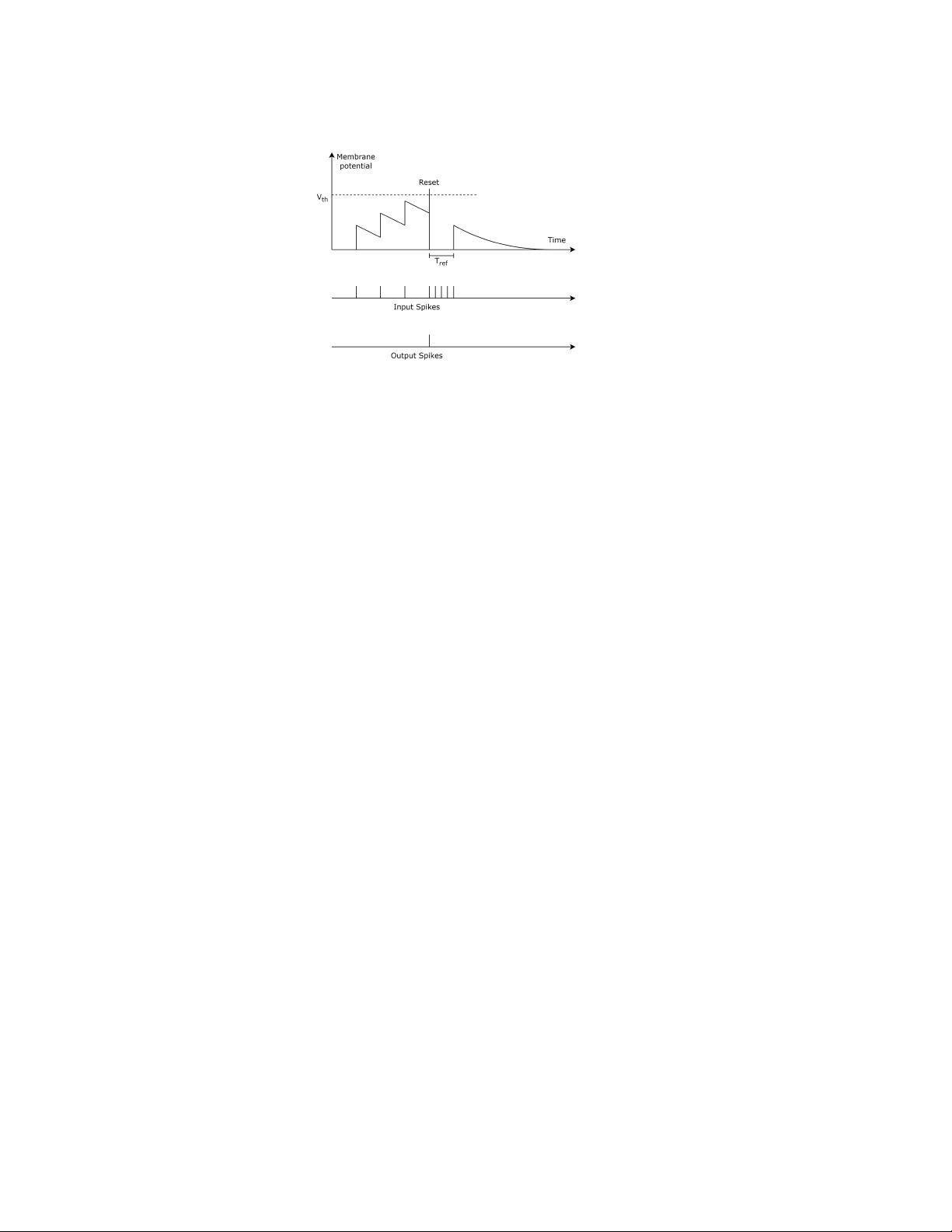
4.1 Spiking Neuron Models and Spike Codes
Spiking Neural Networks (SNNs) are a realistic model of biological networks [
61
,
126
]. While in traditional Articial
Neural Networks (ANNs), neurons communicate via real-valued signals, in SNNs they emit short pulses called spikes.
All the spikes are equal to each other and values are encoded in the timing or in the frequency with which spikes are
emitted.
Various spiking neuron models have been proposed in literature [61], which we highlight in the following:
Manuscript submitted to ACM
剩余30页未读,继续阅读
点击了解资源详情
166 浏览量
118 浏览量
329 浏览量
2023-08-02 上传
153 浏览量
155 浏览量
259 浏览量

学术菜鸟小晨
- 粉丝: 2w+
- 资源: 5753
 我的内容管理
展开
我的内容管理
展开







最新资源
- Web-projekat:Projekat iz predmeta Web程序
- TDD论坛
- noisia:PostgreSQL有害的工作负载生成器
- dgcabkwu.zip_三维数据分析_三维连通域_时域数据图
- Torpedo
- C#MFC串口通信实现
- speedyplane2247csgo.github.io
- TMP117_51.zip
- opengels2.0颜色混合.zip
- WebLogReader网站日志阅读器 v1.0
- 设备方向:用于检测设备方向和运动的Web组件(带有Polymer)
- 安卓Android图书馆座位占座app设计可导入AndroidStudio
- KSEM 2018 proceedings.zip
- ansoft link(1)
- ArcfaceDemo_CSharp:Arcface2.0 的 C# Demo
- asp.net+sqlserver住哪儿酒店预订网站设计基于html5设计
