多组学分析揭示了肠道微生物对医疗卫生的重要性
54 浏览量
更新于2025-01-16
收藏 1.96MB PDF 举报

工程
15
(
2022
)
115
研究
医疗卫生-文章
多组学分析提供了基于肠道微生物群
裴汉
a
,
#
,李丽莎
b
,
#
,王子喜
b
,
#
,林希
c
,
#
,于航
a
,林聪
a
,张正伟
a
,付
杰
a
,彭冉
a
,潘立斌
a
,马树荣
a
,王雪艳
d
,王洪田
d
,王向东
e
,
王燕
a
,
孙金吕
b
,
姜建东
a
,孙
金吕
a
中国医学科学院北京协和医学院药物研究所天然药物活性物质与功能国家重点实验室,北京
100050
b
中国医学科学院北京协和医院过敏性疾病诊治精准医学北京市重点实验室国家免疫与皮肤病临床研究中心变态反应科,中国
c
耳鼻咽喉头颈外科过敏科耳鼻咽喉头颈外科教育部重点实验室
首都医科大学附属北京同仁医院,北京
100005
d
首都医科大学附属北京世纪坛医院变态反应科,北京
100038
e
首都医科大学附属北京同仁医院北京耳鼻喉科研究所北京市重点实验室,北京
100005
阿提奇莱 因福奥
文章历史记录:
2020年12月27日收到
2021
年
2
月
24
日修订
2021年3月29日接受
2021年5月1日网上发售
关键词:
代谢组
肠道菌群干草热
过敏性疾病
肠屏障功能障碍
A B S T R A C T
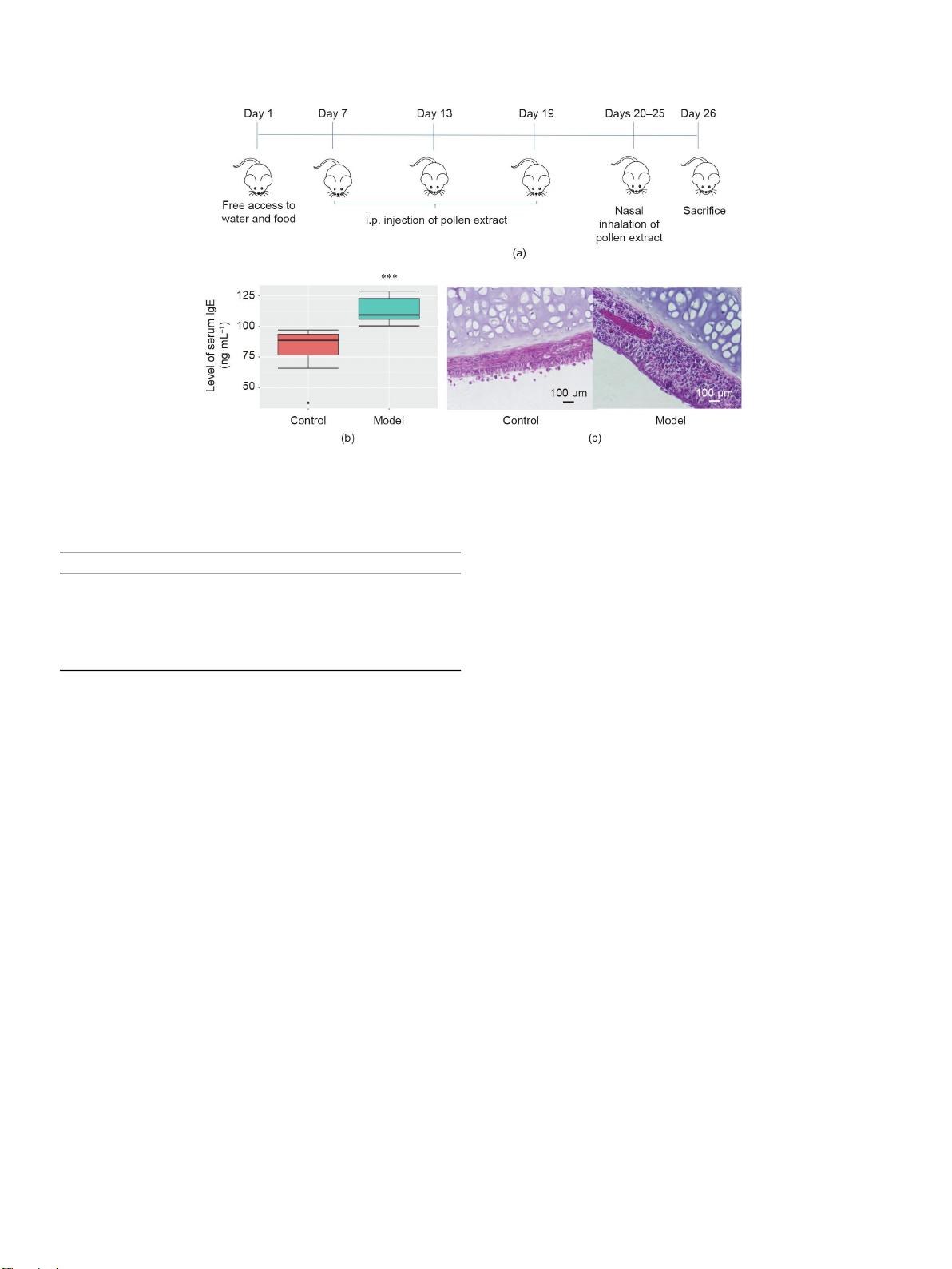
由于变态反应性疾病在世界范围内流行,而且还没有治愈的方法,因此迫切需要探索其病理生理机制。由于过
敏性疾病与肠道生态失调有关,我们从宿主和微生物群之间的分子界面的角度,同时进行代谢组学和微生物群
组成分析,寻找可能的机制。给Sprague-Dawley大鼠注射
蒿属
花粉提取物以刺激对花粉的过度反应。这种过
度反应减少了缬氨酸、异亮氨酸、天冬氨酸、谷氨酸、谷氨酰胺、吲哚丙酸酯(IPA)和肌醇的循环,并减少
了粪便中的短链脂肪酸(SCFA)。几个有益的属属于瘤胃球菌科,毛螺菌科,梭菌下降模型组,而
螺杆菌
和
阿克曼氏菌
只在模型组中表达。模型组大鼠小肠claudin-3和肝脏脂肪酸结合蛋白表达下调,并与代谢改变和细
菌相关
。
我们的研究结果表明,氨基酸及其衍生物(特别是缬氨酸和IPA,这是色氨酸的还原产物),SCFA
和肠道微生物组(特别
是阿克曼氏菌
和
螺杆菌
)的改变可能会通过抑制claudin蛋白的表达和影响粘液层来破坏
肠道屏障功能,这进一步导致花粉热。
©2021 THE COUNTORS.由爱思唯尔有限公司代表中国工程院和高等教育出版社有限公司出版。这是一篇CC
BY-NC-ND许可下的开放获取文章
(
http://creativecommons.org/licenses/by-nc-nd/4.0/
)中找到。
1.
介绍
花粉热是一种季节性过敏性鼻炎(AR),主要影响上呼吸系统。它
是由吸入植物花粉引起的,只发生在特定的个体群体中[1]。对风媒花粉
和虫媒花粉的过敏反应均有报道,症状各不相同。
*
通讯作者。
电 子 邮 件 地 址 :
entwxd@vip.sina.com
(
X.- D. Wang
) ,
wangyan@imm.ac.cn
(
Y.Wang
),
sunjinlv@pumch.cn
(
J.- L. Sun
),
jiang. 163.com
(
J.- D. Jiang
)。
#
这些作者对这项工作做出了同样的
疾病的严重程度取决于个人的具体健康状况[2,3]。迄今为止,全世界
已确认有150多种花粉过敏原来源于草、杂草和树木[4]。在北美和欧
洲,主要致敏花粉分别来自
豚草属
和禾本科。在中国,特别是在北方,
蒿属
和
葎草
属花粉是秋季花粉热的主要贡献者[5,6]。
花 粉 热 病 理 学 的 主 流 学 说 主 要 集 中 在 不 平 衡 的 免 疫 系 统 。
Wambre
等人
[7]
首先发现了
2
型辅助性
T
细胞的一个亚群,命名为
T
H
2A
,这是与过敏性疾病发病机制有关的特异性细胞尽管对过敏性疾
病免疫成分的研究
https://doi.org/10.1016/j.eng.2021.03.013
2095-8099/©2021 THE COMEORS.
由爱思唯尔有限公司代表中国工程院和高等教育出版社有限公司出版。这是一篇基于
CC BY-NC-ND
许可证的开放获取文章
(
http://creativecommons.org/licenses/by-nc-nd/4.0/
)。
可在ScienceDirect上获得目录列表
工程
杂志 首页:
www.elsevier.com/locate/eng

剩余10页未读,继续阅读
172 浏览量
142 浏览量
2021-12-24 上传
143 浏览量
103 浏览量
2021-02-26 上传
106 浏览量
130 浏览量

cpongm
- 粉丝: 6
 我的内容管理
展开
我的内容管理
展开







最新资源
- 全面详实的大学生电工实习报告汇总
- 利用极光推送实现App间的消息传递
- 基于JavaScript的节点天气网站开发教程
- 三星贴片机1+1SMT制程方案详细介绍
- PCA与SVM结合的机器学习分类方法
- 钱能版C++课后习题完整答案解析
- 拼音检索ListView:实现快速拼音排序功能
- 手机mp3音量提升神器:mp3Trim使用指南
- 《自动控制原理第二版》习题答案解析
- 广西移动数据库脚本文件详解
- 谭浩强C语言与C++教材PDF版下载
- 汽车电器及电子技术实验操作手册下载
- 2008通信定额概预算教程:快速入门指南
- 流行的表情打分评论特效:实现QQ风格互动
- 使用Winform实现GDI+图像处理与鼠标交互
- Python环境配置教程:安装Tkinter和TTk
