【进化树构建原理】:10分钟快速理解算法与应用
发布时间: 2025-01-09 00:02:03 阅读量: 5 订阅数: 7 

人工智能实验报告:决策树、循环神经网络、遗传算法、A*算法、归结原理

# 摘要
进化树构建是研究生物进化关系的重要方法,对于分类学、遗传学等领域具有不可或缺的作用。本文首先介绍了进化树的概念及其在生物学研究中的重要性,随后详细阐述了进化树构建的理论基础,包括遗传学与分子生物学原理、不同进化树构建方法及其评价比较。文章进一步探讨了在实践中运用各种进化树构建软件工具所需的数据准备、参数设置以及解读进化树的方法。此外,本文还分析了在复杂数据集和跨学科研究中构建进化树时面临的挑战和可能的解决策略,并展望了未来进化树构建技术的发展方向,强调了人工智能和跨学科研究的重要性。最后,提供了进一步学习进化树构建的资源推荐,以供读者深入研究和实践。
# 关键字
进化树构建;生物进化;理论基础;软件工具;数据处理;解读方法;人工智能;跨学科研究
参考资源链接:[PHYLIP软件详解:系统发育树构建与分子进化分析](https://wenku.csdn.net/doc/ac1udzyj15?spm=1055.2635.3001.10343)
# 1. 进化树构建的概念与重要性
## 1.1 进化树的基本概念
进化树,又称系统发育树,是用来表示物种或基因在进化过程中关系的一种图形化表示方法。它的每个节点代表一个共同祖先,分支的长度通常代表进化的时间距离,而末端分支则代表当前存在的物种或基因。构建进化树是生物学研究中的一项重要任务,因为它们帮助科学家揭示物种的起源、演化过程以及不同物种间的亲缘关系。
## 1.2 进化树构建的重要性
进化树构建的重要性不言而喻。在生物学研究领域,准确的进化树可以为疾病防治、生态管理、物种保护、遗传资源的利用等提供重要的科学依据。通过进化树,研究人员可以确定基因的演化路径,预测疾病的遗传基础,以及理解复杂生物过程的进化机制。此外,进化树在诸如古生物学、流行病学和人类学等领域都发挥着至关重要的作用,使得不同学科的研究者能够从一个共同的视角来审视和理解生命进化的历史。
通过本章内容,读者应能理解进化树在研究中的基础地位和在多个学科中的应用价值,并为后续章节中探索进化树构建的更深层次方法、工具和应用实例打下基础。
# 2. 进化树构建的理论基础
进化树,作为理解生命演化过程的重要工具,不仅体现了物种之间的亲缘关系,还能揭示基因和蛋白质的进化路径。在本章节中,我们将深入了解进化树构建的理论基础,涵盖遗传学与分子生物学的进化原理、进化树的构建方法,以及如何评价与比较不同的进化树。
### 2.1 遗传学与分子生物学的进化原理
在进化生物学中,DNA序列的变异是遗传多样性产生的基础,同时也是研究物种演化关系的核心。分子钟假说与物种分化时间估计则是推断物种演化历史的重要理论依据。
#### 2.1.1 DNA序列的变异与遗传多样性
DNA序列的变异通常分为两种:点突变和大范围的结构变异。点突变是指单个核苷酸的替换,而结构变异则涉及大片段DNA的插入、缺失、复制或重组。这些变异在物种进化过程中积累,形成了遗传多样性。我们可以通过比较不同物种的DNA序列来发现这些变异,进而研究其演化关系。
#### 2.1.2 分子钟假说与物种分化时间估计
分子钟假说提出,DNA序列的替换速率在不同物种间大致保持恒定,因此可以用该速率来估计物种分化的时间。根据分子钟假说,通过比较两个物种的特定DNA序列差异,可以计算出它们自分离以来的相对时间。尽管这个理论在实际情况中会有所偏差,它仍然是一个强大的工具,用于研究物种的历史演化。
### 2.2 进化树的构建方法概述
进化树的构建方法有多种,主要包括最大简约法(MP)、最大似然法(ML)和贝叶斯推断(BI)。每种方法都有其优势和局限性,研究者可根据数据特点和研究目的选择合适的方法。
#### 2.2.1 最大简约法(MP)
最大简约法是最早应用于系统发育分析的方法之一。它基于这样的假设:进化过程中,使特征数最少的树(即简约树)最可能代表实际发生的进化关系。MP方法不考虑序列的进化速率,因此在处理含有大量简约信息的数据集时更为有效。
#### 2.2.2 最大似然法(ML)
最大似然法是一种统计学方法,它寻找在已知模型下得到观测数据概率最大的树。它需要先设定一个进化模型,然后基于此模型估算树的分支长度和拓扑结构,最终选出最有可能产生观测数据的树。ML方法能够提供关于进化树分支置信度的统计估计。
#### 2.2.3 贝叶斯推断(BI)
贝叶斯推断是一种基于概率的模型选择方法。它通过对可能的进化树进行概率分配,并在观测数据下调整这些概率,最后根据贝叶斯定理得到后验概率最高的进化树。贝叶斯推断因其能够处理不确定性并整合先验知识而变得越来越流行。
### 2.3 进化树的评价与比较
构建多个进化树后,需要对它们进行评价与比较,以确认哪棵树最能准确反映物种间的演化关系。树的置信度和重聚概率是评估进化树质量的重要指标。
#### 2.3.1 树的置信度和重聚概率
树的置信度通常通过重聚概率(如自举检验值,即bootstrap值)来评价。重聚概率是指在从原始数据中通过重复抽样得到的许多新数据集上,某个节点在进化树上被重新发现的频率。一个高重聚概率的节点表明它具有较高的可靠性。
#### 2.3.2 树的比较方法与统计显著性测试
比较不同的进化树时,可以使用各种统计方法来评估它们之间的一致性。例如,使用Kishino-Hasegawa(KH)测试、Shimodaira-Hasegawa(SH)测试或贝叶斯因子测试等方法来检验不同的进化树之间的统计显著性差异。这些测试有助于研究者选择最符合数据的进化树。
在下一章节,我们将介绍实际使用中的进化树构建软件工具,并展示如何利用这些工具进行进化树的构建实践。
# 3. 进化树构建的软件工具实践
在现代生物信息学中,进化树的构建不再是一项繁重的手动任务,而是通过各种专门的软件工具来实现自动化、高效率的分析。本章节将详细介绍几种常用的进化树构建软件,讨论如何准备数据以及设置参数,以构建准确的进化树。
## 3.1 常用进化树构建软件介绍
### 3.1.1 MEGA与进化树的绘制
MEGA(Molecular Evolutionary Genetics Analysis)是一款流行且用户友好的进化分析软件。它不仅适用于进化树的绘制,还能进行序列分析和多样性分析。MEGA提供了一个可视化的界面,方便用户导入、编辑和分析序列数据,以及进行进化树的构建和修饰。
#### 主要特点:
- **用户界面友好**:适合初学者和有经验的科学家。
- **多种进化树构建方法**:支持NJ(邻接法)、MP(最大简约法)、ML(最大似然法)等多种算法。
- **统计检验**:提供Bootstrap测试来评估树的置信度。
#### 操作步骤:
1. **序列导入**:通过文件菜单导入或直接拖放序列文件到MEGA界面。
2. **序列对齐**:使用ClustalW、MUSCLE等内置工具进行序列对齐。
3. **进化树构建**:选择合适的进化树构建方法,输入相应的参数,然后执行构建命令。
4. **树的可视化与编辑**:绘制的进化树可以进行修饰,如分支颜色、标签等,并导出为图片。
### 3.1.2 MrBayes与贝叶斯进化分析
MrBayes是一款专门用于贝叶斯进化分析的软件。它通过模拟Markov链Monte Carlo(MCMC)来估计后验概率分布,提供了一种比传统方法更加复杂和灵活的进化树构建方式。
#### 主要特点:
- **贝叶斯方法**:基于概率模型构建进化树,能考虑进化过程中的不确定性。
- **高级的MCMC技术**:可以模拟复杂的进化模型,评估模型参数。
- **并行处理**:支持多线程计算,能显著提高计算速度。
#### 操作步骤:
1. **配置文件准备**:根据MrBayes的语法编写配置文件,定义进化模型、参数等。
2. **命令行运行**:在终端或命令提示符中执行MrBayes命令,输入配置文件。
3. **结果分析**:分析输出文件,评估模型收敛性和树的后验概率。
4. **树图生成**:根据结果文件生成进化树图。
### 3.1.3 PHYLIP与多种算法的实现
PHYLIP(PHYLogeny Inference Package)是一个功能强大的命令行工具包,提供了多个不同算法来构建进化树,包括距离法(如NJ)、最大似然法和最大简约法等。
#### 主要特点:
- **命令行操作**:适合在服务器或终端中使用。
- **算法多样性**:支持多种进化树构建算法。
- **高效的计算性能**:适用于大型数据集的分析。
#### 操作步骤:
1. **安装PHYLIP**:根据操作系统下载并安装PHYLIP。
2. **准备输入文件**:通常为序列对齐后的文件。
3. **命令执行**:使用不同的PHYLIP命令(如`protdist`、`dnapars`等)来执行进化树的构建。
4. **结果查看**:分析输出文件,如`*.out`文件,其中包含了进化树的结果。
## 3.2 数据准备与格式转换
### 3.2.1 序列的对齐与预处理
进化树构建的准确性在很大程度上依赖于输入数据的质量,尤其是序列的对齐质量。序列对齐是将来自不同物种或个体的DNA、RNA或蛋白质序列按照相似性排列在一起的过程。不正确或不一致的对齐会对最终的树造成重大误差。
#### 操作流程:
1. **序列选择**:从数据库如NCBI或UniProt中获取相关物种或个体的序列。
2. **对齐工具选择**:使用ClustalW、MUSCLE等对序列进行初步对齐。
3. **对齐结果检查与修正**:检查并手动修正对齐结果,尤其是重要功能区段的对齐。
### 3.2.2 文件格式的转换与兼容性问题
不同的进化树构建软件通常需要特定的文件格式。因此,数据格式转换是一个常见的需求,确保数据能被所选软件正确读取和分析。
#### 主要格式:
- **FASTA**:最常用的序列格式,以">"标识符开头,后面跟着序列信息。
- **PHYLIP**:一种广泛使用的格式,通常用于PHYLIP软件包。
- **Nexus**:一个较为复杂的格式,常用于贝叶斯进化分析软件,如MrBayes。
#### 文件格式转换工具:
- **Seqotron**:适用于多种格式之间的转换。
- **EMBOSS**:一组丰富的命令行工具,支持序列格式转换。
- **BioEdit**:图形用户界面程序,简化了多种序列格式的转换。
## 3.3 进化树的构建步骤与参数设置
### 3.3.1 进化模型的选择与适用性
进化模型是构建进化树的重要参数。它决定了序列变化的速率和模式,不同的模型适用于不同的数据和进化场景。
#### 常见进化模型:
- **Jukes-Cantor (JC) 模型**:一个最简单的模型,假设所有位点的替换率相同。
- **Kimura模型**:区分了转换(transition)和颠换(transversion)的速率。
- **GTR模型**(General Time Reversible):最灵活的模型之一,可以估计不同碱基间的转换速率。
### 3.3.2 运行构建进化树的命令与界面操作
选择适当的进化模型后,我们需要设置合适的命令和参数以运行进化树构建过程。在命令行工具(如MEGA)中,这一步通常通过图形界面进行设置。
#### 运行示例:
```bash
# 使用MEGA的命令行版本MEGAX进行操作
megax --phylogeny --data_file myalignedsequences.fasta --model JC --out mytree.nwk
```
在上述命令中,我们指定了进化模型为JC,数据文件为`myalignedsequences.fasta`,输出文件为`mytree.nwk`。
```mermaid
graph TD
A[开始构建进化树] --> B[选择合适的进化模型]
B --> C[准备输入文件]
C --> D[配置并运行软件]
D --> E[评估进化树的置信度]
E --> F[树的优化与调整]
F --> G[结束]
```
#### 参数设置:
- **数据文件**:对齐后的序列数据。
- **进化模型**:根据数据特点选择合适的模型。
- **参数估计**:如进化率、树分支长度等。
- **统计测试**:如Bootstrap或自举方法。
在进行参数设置时,需要综合考虑计算资源、数据集大小和复杂性。对于大数据集,可能需要额外的优化设置来保证计算的高效性。
本章节介绍了进化树构建的软件工具,并详细讲述了数据准备、序列对齐、格式转换以及关键的进化模型和参数设置。通过掌握这些基础知识,研究者可以更准确地构建出能够反映物种间真实进化关系的进化树。
# 4. 进化树的解读与应用实例
### 4.1 进化树的解读方法
进化树是展示物种进化关系的一种图形化模型。正确解读进化树对于理解物种间的演化历程至关重要。本节将介绍进化树的基本阅读方式以及如何解读其中的关键信息。
#### 4.1.1 树的阅读与节点解释
进化树由节点和分支组成,其中节点表示物种、亚种或其他分类群,分支表示物种间的进化关系。一个进化树的根节点代表着共同的祖先,从根节点延伸出的分支可以连接到其他节点,展示物种随时间的演化路径。
解读进化树时,首要关注的是分支长度和分支之间的角度。通常,分支的长度与遗传距离成正比,即两个物种的DNA序列差异越大,它们之间的分支就越长。分支间的角度大小并不重要,因为进化树通常可以进行旋转而不改变树中的信息。
#### 4.1.2 物种关系的推断与比较
解读进化树的第二个关键在于理解物种之间的关系。通过比较不同分支的连接方式,我们可以推断物种间的亲缘关系。例如,如果两个物种的分支从一个共同的节点延伸出来,那么这两个物种有较近的演化关系。
在比较进化树时,可能需要注意以下几点:
- 树的拓扑结构,即分支的连接方式,能够反映物种的亲缘关系。
- 支持度值,如自展值(bootstrap value),显示了节点稳定性的统计度量。通常,支持度值越大,该节点表示的关系就越可靠。
- 演化时间的估计,通过分子钟假设,可以大致推断出物种分化的时间。
### 4.2 进化树在分类学的应用
进化树在分类学中是一个非常有用的工具,用于确定物种的分类地位以及构建更为精确的分类体系。
#### 4.2.1 物种鉴定与分类地位的确定
进化树可以帮助生物学家鉴定未知物种。通过将未知物种的基因序列与已知物种的序列进行比较,并构建进化树,科学家可以推断出未知物种的分类地位。
例如,一个新发现的鸟类物种的DNA序列可以被用来构建一个进化树,与其他鸟类物种进行比较。如果该物种的基因序列与蓝鸟非常相似,那么可以推断出它与蓝鸟属于近亲,进而有助于确定其分类地位。
#### 4.2.2 进化关系与分类体系的构建
进化树不仅能用于单个物种的鉴定,还能帮助构建整个生物群组的分类体系。通过分析多个物种之间的基因序列差异和演化关系,科学家能够更好地理解物种间的亲缘关系,并据此调整分类体系。
例如,使用分子系统发育分析,生物学家们可能发现某些传统的分类方法并没有正确反映物种的进化关系。基于进化树的分析,科学家们可以重新定义物种的分类界限,使之更符合演化事实。
### 4.3 进化树在遗传学的研究
除了分类学之外,进化树也是遗传学研究中不可或缺的工具,用于探索基因的演化历史和遗传疾病的进化机制。
#### 4.3.1 基因的演化历史研究
进化树能够展现特定基因的演化历史。研究者可以构建包含不同物种的基因序列的进化树,以研究这些基因是如何随物种分化而变化的。
例如,在研究抗药性基因时,科学家可以追踪这些基因在不同细菌谱系中的出现时间和演化路径。这有助于了解抗药性基因的传播机制,并为控制抗药性的发展提供策略。
#### 4.3.2 疾病遗传机制的进化分析
进化树还可以帮助遗传学家研究某些遗传疾病的起源和传播。通过分析遗传疾病的基因在进化树上的位置,科学家可以了解这些基因的起源时间,并推测其在人群中传播的可能途径。
例如,在研究遗传性疾病的进化历史时,研究者可能发现某些基因突变是相对古老的,并且在多个物种中均有出现。这提示我们这些基因突变可能在物种分化之前就已经存在,并对物种的生存有着重要的影响。
```mermaid
graph TD
A[物种A] -->|进化| B[物种B]
A -->|进化| C[物种C]
C -->|进化| D[物种D]
D -->|进化| E[物种E]
```
```markdown
在上述Mermaid流程图中,我们展示了物种间进化的关系,从物种A逐步分化出物种B、C,直至物种E的演化路径。
```
通过本章的学习,你应该已经对进化树的解读方法以及进化树在分类学和遗传学中的应用有了深入的理解。在后续章节中,我们将进一步探讨构建复杂进化树的挑战、进化树与其他生物信息学工具的整合,以及进化树未来的发展方向。
# 5. 进化树构建的高级主题
随着生物信息学的快速发展,进化树构建技术也面临着越来越多的挑战与机遇。在这一章节中,我们将探索更复杂的进化树构建问题,探讨进化树与其他生物信息学工具的整合方式,并展望进化树构建技术未来的发展方向。
## 5.1 复杂进化树的构建挑战
随着基因组学和转录组学数据的日益丰富,进化树构建面临着更多的挑战,尤其是在处理复杂进化事件和大数据集方面。研究者需要解决的关键问题包括并行演化和隐性基因流,以及大数据集带来的计算挑战。
### 5.1.1 并行演化与隐性基因流问题
并行演化是指不同物种独立演化出相似的特征。这种情况在进化树中表现为多个分支的相同分支节点,可能会导致错误的物种间关系推断。研究者需要使用更复杂的模型来区分真正的同源特征与并行演化产生的相似特征。
隐性基因流则是指物种间的基因交换,这可能干扰进化树的构建,因为基因流会导致遗传信息在物种之间共享。目前,已经有新的统计模型和算法被提出,用于检测和校正隐性基因流的影响,如树混合模型(TreeMix)。
### 5.1.2 大数据集的计算挑战与优化
现代生物信息学研究产生的数据量巨大,这对进化树构建的计算效率提出了更高的要求。为了处理大规模数据集,研究者们开发了多种优化策略。一种常见的做法是使用并行计算技术,比如分布式计算框架(如Apache Hadoop或Apache Spark)来加速进化树构建过程。
此外,算法的优化也是提高计算效率的关键。例如,RAxML是一种广泛使用的进化树构建软件,它通过使用快速的启发式搜索策略和高效的并行实现,能够处理大规模数据集的进化树构建问题。
### 代码块:使用RAxML进行进化树构建的示例
以下是一个使用RAxML软件进行进化树构建的基本示例:
```bash
# 安装并调用RAxML
module load raxml
raxml -f a -m GTRGAMMA -s data.phy -n output
```
在这个命令中:
- `-f a` 表示运行快速自举和最优化搜索的组合。
- `-m GTRGAMMA` 指定了进化模型,即通用时间可逆模型加上伽马分布速率变异。
- `-s data.phy` 是输入文件,包含需要构建树的多序列比对。
- `-n output` 指定输出文件的基本名。
### 5.2 进化树与其他生物信息学工具的整合
进化树构建不是孤立的,它需要与其它生物信息学工具和数据源整合,以获得更全面的生物信息学分析结果。
### 5.2.1 基因组学与转录组学数据的整合分析
基因组学和转录组学的数据提供了关于基因和它们表达的信息,这些数据可以与进化树相结合,以探索特定基因家族在进化过程中的变化。
例如,通过比较不同物种的基因组,研究者可以识别出在特定物种中扩展或收缩的基因家族,并使用进化树来推断这些变化发生的时间点和可能的原因。
### 5.2.2 多组学数据与系统生物学的关系构建
多组学数据整合了基因组、转录组、蛋白质组、代谢组等多个层面的生物信息。通过将这些数据与进化树结合起来,研究者能够构建更为复杂的生物学系统模型。
在多组学数据整合的过程中,进化树作为参考框架,帮助研究者理解在不同的环境压力或生理条件下,基因及其表达的变化是如何适应和进化的。
### 5.3 进化树未来的发展方向
随着计算能力的提升和新的生物信息学方法的发展,进化树构建技术也在不断地进步。
### 5.3.1 人工智能在进化树构建中的应用前景
人工智能特别是机器学习技术的引入,为进化树构建带来了新的可能。这些技术可以用于识别复杂的进化模式,比如通过深度学习预测蛋白质结构,并在此基础上推断进化关系。
此外,人工智能可以处理和分析大量的数据集,提供更精准的进化关系预测,并能够自动化执行复杂的进化树构建流程,提高效率和准确性。
### 5.3.2 跨学科研究在进化树分析中的作用
进化树构建是一个跨学科的领域,涉及生物学、数学、计算机科学等多个学科。未来的进化树构建将更加依赖于跨学科团队的合作。
例如,数学模型的改进可以提供更精确的进化分支预测;而计算机科学的进步则能够帮助处理更大的数据集,并将进化树与系统生物学模型相结合。
在本章节中,我们探讨了构建复杂进化树所面临的挑战,进化树与其他生物信息学工具的整合,以及未来的发展方向。通过理解和应用这些高级主题,研究者可以更深入地探索生物多样性的演化历史,并在更广阔的生物学背景下理解生命的进化。
为了进一步探索这个主题,下一章节将总结进化树构建的实践意义,并推荐一些学习资源,以供有兴趣深入了解该领域的读者参考。
# 6. 结语与学习资源推荐
进化树构建不仅仅是一个理论问题,它对于生物科学的多个分支领域都具有深远的实践意义。本章节将总结进化树构建的实践意义,并推荐一些进一步学习进化树构建的资源。
## 6.1 进化树构建的实践意义总结
进化树构建是生物信息学领域的基础,它帮助研究者描绘出物种之间的亲缘关系。在生物分类学中,进化树有助于明确物种的分类地位,推动新的分类体系的建立。在医学领域,通过研究基因的演化历史,科学家可以更好地理解疾病的遗传机制,为疾病预防和治疗提供新的思路。此外,进化树还在环境科学、农业研究、微生物生态学等领域发挥着重要作用。
## 6.2 进一步学习进化树构建的资源
### 6.2.1 在线课程与教程
为了深入学习进化树构建,您可以参考以下在线资源:
- Coursera 提供的“生物信息学入门”课程,其中涉及进化树的构建与分析。
- edX 平台上的“高级计算生物学”系列课程,涵盖了进化树构建的高级主题。
- Bioinformatics.ca 提供的一系列专业教程,尤其适合对软件工具实践感兴趣的学习者。
### 6.2.2 学术论文与最新研究成果
要跟上进化树构建的最新研究进展,以下是一些重要的学术资源:
- Nature Genetics 和 PNAS 等顶级科学期刊,定期发表关于进化树构建的最新研究。
- Google Scholar 和 PubMed 是搜索相关学术论文的强大工具,可利用关键词“evolutionary tree construction”进行检索。
- “Molecular Biology and Evolution”杂志是专注于分子生物学和进化研究的重要出版物。
### 6.2.3 开源工具与社区交流平台
开源工具和社区平台为学习和实践提供了良好的环境:
- GitHub 上有多个开源进化树构建项目的仓库,如BEAST和RAxML,可以下载源代码进行深入研究。
- BioStars 是一个生物信息学问题和解答的社区,可以在这里提问和交流。
- The Open Tree of Life 项目旨在构建和展示生命全树,其网站上提供了丰富的进化树数据和相关资源。
通过对进化树构建理论和实践的深入理解,并利用上述推荐资源,无论是初学者还是有经验的研究者都可以在这一领域不断进步和探索。随着计算能力的提高和数据分析技术的发展,进化树构建将不断拓展其应用范围,为生物科学贡献更多的洞见。

 百万级
高质量VIP文章无限畅学
百万级
高质量VIP文章无限畅学
 千万级
优质资源任意下载
千万级
优质资源任意下载
 C知道
免费提问 ( 生成式Al产品 )
C知道
免费提问 ( 生成式Al产品 )
0
0
相关推荐

专栏简介
《进化树分析:终极指南》专栏提供了一系列全面的文章,涵盖了系统发育树分析的各个方面。从入门技巧到高级技术,该专栏指导读者掌握系统发育树软件,提升研究效率。文章深入探讨了进化树构建原理、数据准备技巧、可视化方法、验证技巧和应对复杂数据的策略。此外,专栏还提供了实战案例,从数据收集到结论得出,全面展示了系统发育树分析的完整流程。无论您是刚接触系统发育树分析的新手,还是经验丰富的研究人员,该专栏都能为您提供宝贵的见解和实用技巧,帮助您提升研究成果。
专栏目录

文章持续更新中,敬请期待~
最低0.47元/天 解锁专栏
买1年送3月
百万级
高质量VIP文章无限畅学
千万级
优质资源任意下载
C知道
免费提问 ( 生成式Al产品 )
最新推荐
【TVbox v4.3.3:技术演进揭秘】

# 摘要
随着数字化时代的加速发展,智能电视盒子市场需求持续增长。本文深入探讨了TVbox v4.3.3的技术背景、市场需求和核心技术,包括其系统架构、性能优化以及安全机制等关键方面。文章还详细分析了TVbox v4.3.3的开发实践,面临的挑战以及如何解决这些挑战。此外,本文强调了TVbox v4.3.3在功能亮点和
深入挖掘FontCreator:个性化字体设计的6个高级技巧

# 摘要
本文详细介绍了FontCreator软件,包括其功能概述及个性化字体设计的理论与实践应用。首先,阐述了字体设计的基本原则和版权合法性,为理解字体设计提供了理论基础。接着,深入探讨了FontCreator中高级编辑技巧,如曲线编辑、字符变体和自动化设计流程的应用。实践应用章节则聚焦于如何创造独特的字体风格、扩展和管理字体家族以及实现字体的跨平台适配。最后,
【树莓派Dlib终极安装指南】:2023年新手必读,一步到位解决所有问题

# 摘要
树莓派作为一种低成本、功能强大的单板计算机,与Dlib这一高效的人工智能库结合使用,开启了一系列激动人心的应用。本文首先介绍了树莓派和Dlib的基本概念,然后详细阐述了树莓派系统的设置与优化,包括操作系统的选择、性能调整、系统安
【公式树方法深入】:高级技巧与策略提升逻辑证明效率

# 摘要
公式树方法作为一种结构化表示和处理逻辑表达式的技术,在逻辑证明、人工智能、软件工程等众多领域中发挥着重要作用。本文首先概述了公式树的概念和理论基础,阐述了公式树的定义、结构以及在逻辑证明中的应用。接着,探讨了构建高效公式树的策略,包括树结构的选择、剪枝和优化技术以及算法性能提升方法。文章还详细介绍了公式树的实践应用和高级技术,如多维结构构建、动态公式树的实时证明及并行化处理。最后,
状态空间分析】:对角线化方法,构建控制系统的理论基石
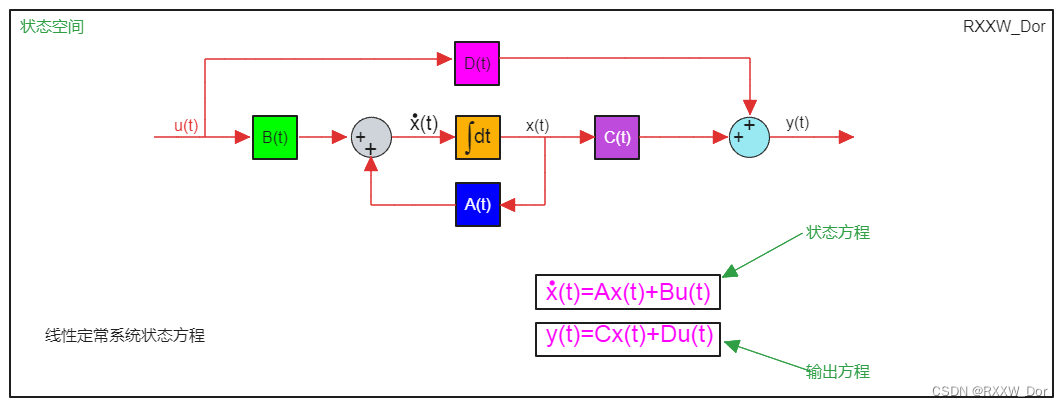
# 摘要
本文系统地介绍了对角线化方法的理论基础及其在控制系统中的应用。首先,概述了对角线化的概念、状态空间表达式以及控制系统的类型和稳定性判定。随后,深入探讨了线性代数中的对角化条件与过程,以及状态空间模型的对角线化步骤和系统解耦。文章进一步阐述了对角线化技术在控制系统设计和分析中的实际应用,包括系统设计步骤、方法优势与局限性,以及系统性能分析和优化案例。高级主题涵盖了非线性系统的线性化和多变量系统的对
微信小程序音频播放技巧:揭秘流畅背后,提升用户体验的播放秘籍
# 摘要
本文全面探讨了微信小程序中音频播放的技术基础、实践技巧与性能优化。首先介绍了音频播放的基础知识,包括音频格式和编解码技术。其次,通过解析微信小程序对音频格式的支持和音频播放器的实现方法,阐述了音频流式传输技术和播放控制的优化策略。接着,文章深入分析了音频播放性能的优化方法,包括缓存策略、内存管理以及异步加载和预加载技术。最后,通过案例分析总结了微信小程序音频播放的成功经验和未来发展趋势
Visual DSD入门秘诀:5分钟构建高效企业数据模型
# 摘要
本文全面解析了Visual DSD这一数据模型设计工具的概念、基础操作和高级应用。文章首先介绍了Visual DSD的基本概念及其安装和配置过程,然后详细阐述了如何创建和编辑数据模型,包括设计模型结构、添加实体和属性,以及使用图形化元素进行模型优化。随后,本文探讨了在实际应用中实现复杂实体关系、集成业务规则和进行版本控制与团队协作的高级应用。通过企业级数据模型的构建和性能优化案例分析,本文分享了实践中的经验教训和最佳实践。最
【MC20901-V1_08.pdf安全特性揭秘】:企业级应用的不二守护神
# 摘要
本论文旨在探讨MC20901-V1_08.pdf的安全特性及其理论基础和实际应用。首先,文章阐述了安全特性的核心概念,包括加密技术、认证和授权机制,以及密钥管理与交换技术。其次,文章深入分析了安全特性的实现技术,如审计日志、访问控制策略和数据完整性维护。进一步,论文通过企业级应
DRYPIX 3500用户反馈解读:5个核心问题与针对性改进方案
# 摘要
本文对DRYPIX 3500产品进行了全面的分析,从用户反馈的核心问题入手,系统地探讨了用户体验、设备性能以及客户服务方面存在的问题,并提出了针对性的改进方案。通过理论探讨和实践应用案例分析,文章详细阐述了提升用户界面易用性、增加设备响应速度和稳定性、以及优化客户服务流程的具体策略。通过实施这些改进方案,DRYPIX 3500在用户满意度、设备性能和服务质量方面均有所提升。最后,本文对未来的改进方向和用户满意度提升策略进行了展望。
# 关键字
DRYPIX 3500;用户体验;设备性能;客户服务;改进方案;满意度提升
参考资源链接:[DRYPIX 3500医用干式成像仪操作手册与
资源上传下载、课程学习等过程中有任何疑问或建议,欢迎提出宝贵意见哦~我们会及时处理!
点击此处反馈


专栏目录
文章持续更新中,敬请期待~
最低0.47元/天 解锁专栏
买1年送3月
百万级
高质量VIP文章无限畅学
千万级
优质资源任意下载
C知道
免费提问 ( 生成式Al产品 )